Summary
Professor: Part 1 of this post is about the beginning of the time course for symptom awareness: Within about .015 seconds, less than one breath, the Saliency Network (SN) of our brain can NOTICE something is strange.
Student: At the end of the third of six stages of relatively fast cortical processing, for example, becoming aware of inspiratory load, we will be sending an alert to the Central Executive Network and dampening the Default Mode Network..
Fast Processing
Student: How quickly do we make critical adjustments before we are ‘done in’?
Professor: These adjustments occur within a breath. In fact, both Gozal et al. (1995) and Raux et al. (2013) compared effects of single versus continuous inspiratory loads.
- WITHIN ONE BREATH, Raux et al. showed:
- increased activity in:
- insula cortex, the hub of interception integration,
- thalamus, the final ‘gate’ to higher centers, and other areas, and
- decreased activity in:
- cingulate cortex,
- temporal-occipital junction, and other areas.
- increased activity in:
Analyzing individual, subcortical, MRI frames was done by Gozal et al. who found:
- AN IMMEDIATE INCREASE in activity of the:
- parabrachial (Vth motor) nucleus, locus coeruleus,
- thalamus, putamen, cerebellum (cumen, central vermis, tuber, & uvula), and other areas
- Post respiratory load recovery followed two time courses:
- immediate decrease, i.e., putamen and cerebellar uvula, or
- slow signal decrease, i.e., basal forebrain and cerebellar vermis.
- Both studies showed decreasing higher CNS activity for continuous load versus single trial of loading, about which Raux et al. suggested this possibibly shows “cortical automatization secondary to motor learning.”
Student: In fact, the charts are very reminiscent of the blog posts you wrote about the adaptation to split-belt walking, especially the involvement of the cerebellum. For examples and discussion, see:
- Control-of-leg-timing-vs-placement-during-split-belt-walking, and
- How-does-the-motor-system-correct-walking-errors?
Professor: Very intriguing recollection of yours about the cerebellum, split-belt walking, and adaptation. Actually, the effect of inspiratory load happens within milliseconds:
- Nf (latency: 25 – 50 milliseconds; f = frontal; possible source in the frontal cortex),
- P1 (latency: 45 – 65 milliseconds; possible source in the centroparietal cortex ),
- N1 (latency:85 – 125 milliseconds; possible source in the in the sensorimotor and frontal cortices)
- P2 (latency: 160 – 230 milliseconds; possible source in the in the sensorimotor and frontal cortices), and
- P3 (latency: 250 – 350 milliseconds; possible source in the parietal cortex)
Professor: A picture is worth a thousand words, so the experts say. Shown below is Figure 3 from the excellent and timely review of von Leopoldt, Chan, Esser, & Davenport (2013) and NOTE THAT NEGATIVE IS UP:
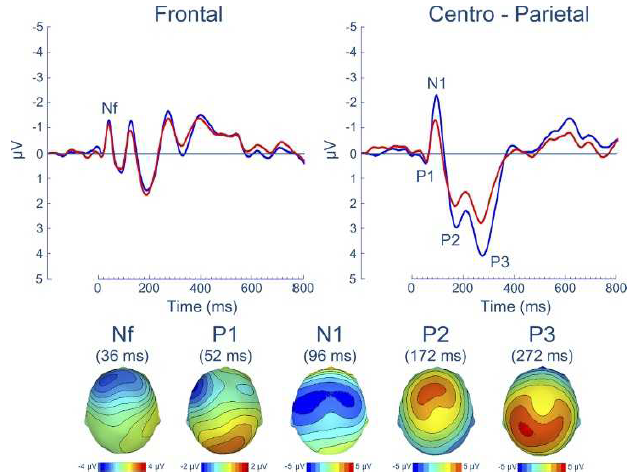
Professor: In Figure 3 are shown the group means for the respiratory-related evoked potential (upper panel) and related scalp topographies at their peak latencies (lower panel), at frontal and centro-parietal regions.
Professor: Also, advanced studies using evoked potentials have identified that most peaks are a composite of different sub-peaks.
Student: Since there are 1,000 ms to a second, all this take place in less than ONE HALF A SECOND.
Professor: And that half a second can be divided into five stages and another stage can be added for producing a subjective report, with the sixth stage taking about another second or so. The following figure illustrate two schemes for the structures and pathways involved:
Schemes
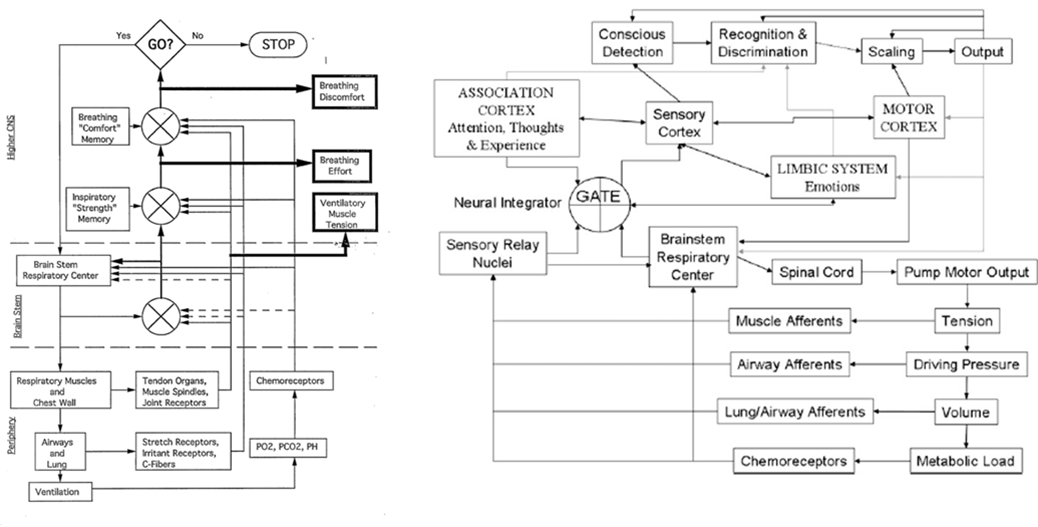
Professor:
- Left figure is from Weiser et al. (1993); notice the three (3)comparators of actual and efference copies, shown as circles with crosses. The resulting three error signals can be self-reported: first, as Ventilatory Muscle Tension; second, as Breathing Effort; third, as Breathing Discomfort.
- Right figure is from Davenport’s contribution to the O’Donnell et al. review (2007). Note the comparator labeled GATE corresponds roughly to the Weiser et al. (1993) comparator for Breathing Effort subjective report.
In six stages
Professor: Below is a model of the time course for sensation processing shown by Figure 5 of Menon & Uddin (2010) with their five stages for processing:
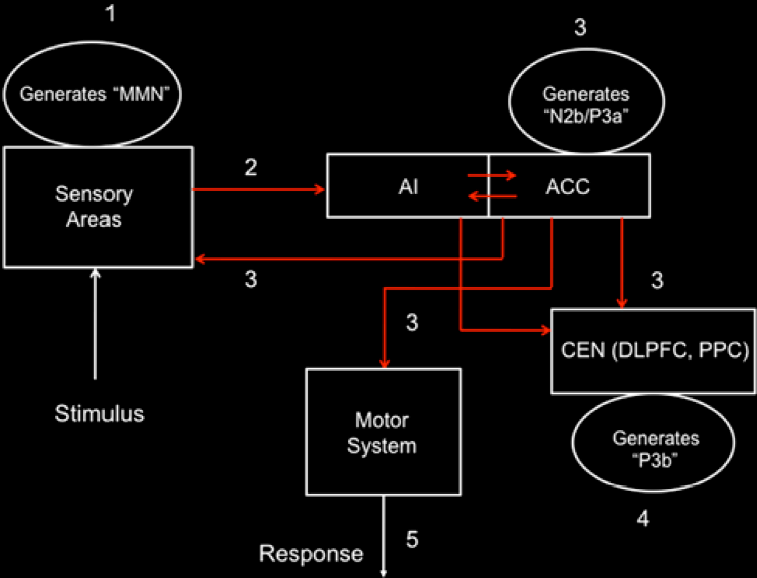
Student: I notice the anterior insula (AI) is tightly connected to the anterior cingulate cortex (ACC), that then influences the central executive network (CEN) composed of the dorsolateral prefrontal cortex (DLPFC) and posterior parietal cortex (PPC). Hmm, “N2b/P3a” is part of the N2 and P3 composite wave and P3b is another part of the P3 wave.
Professor: The numbers in their Figure 5 are the stages of processing. The following are the details for Stage 1:
Stage 1 – At about 150 ms post-stimulus, detection of a ‘deviant’ stimulus by the primary sensory areas, including posterior insula, is indexed by the mismatch negativity (MMN) component of the evoked potential.
Professor: The stimulus is inspiratory loading, frequently done by a brief inspiratory occlusion, which is a ‘deviant’ stimulus:
- Phrenic nerve activity from pulmonary mechanoreceptor and stretch receptors to
- Cervical spinal cord* and cuneate nuclei*
- Cerebellum – to lobule V including intermediate cortex* and vermis* via mossy fibers to ventroposteriolateral nuclei and/or
- Thalamus – to ventroposteriolateral nuclei* from cuneothalamic to fastigal nuclei* to the
- Thalamic reticular nuclei (TRN)* from the ventroposteriolateral thalamic nuclei and
- FINALLY TO THE Posterior insula (PI)*.
Professor: For more on efference copies, see my earlier post: Signals-of-efference-copy-and-effort.
Student: Then those asterisks (*) are the possible ‘efference copy comparator(s)’ and/or ‘MMN source(s)’ for
- ventilatory muscle tension,
- breathing effort, and
- breathing discomfort.
Professor: More on ‘efference copy comparators in the coming post on Perceived Exertion. but here are the details for Stage 2:
Stage 2 – Next is a “bottoms-up” MMN transmission to other brain regions, especially anterior insula (AI) and amygdala. AI provides selective amplification of critical, i.e., salient, events that trigger a strong response in the anterior cingulate cortex (ACC).
Professor: This is not too surprising for MMN transmission to quickly go to AI, since the PI and AI are so tightly connected.
- Together, AI and ACC are the critical areas and form the foundation of the Saliency Network (SN).
- In fact, some ‘blocks’ of the AI “play the role of hubs, bridging the anterior and posterior circuits of the insula” (Cauda et al., 2011).
- In addition, both the PI and AI contain “modality-specific primary sensory representations of each of the affective, interoceptive feelings from the body, and that each representation is organized somatotopically in the anteroposterior direction.” (Craig, 2010).
Student: There is also evidence that PI is interceptively oriented, whereas the AI is organized for integration and decision-making (Cauda et al., 2011).
Professor: Onward and outward throughout the cortex. Here are the details for Stage 3:
Stage 3 –At about 200–300 ms post-stimulus, the AI and ACC will have generated a “top–down” control signal, as indexed by the N2b/P3a component of the evoked potential. This signal is simultaneously transmitted to the primary sensory and association cortices, as well as to the central executive network (CEN).
Professor: The alpha traveling wave with the posterior insula’s searchlight finds a focal disturbance and activates the CEN to switch off/reduce the activity of default mode network (DMN).
Student: Oh, I remember! The Saliency Network (see Oops-I-forgot-and-I-must-reconnect-to-the-breath), or the Tonic Alertness Network (see Windshield-wiping-of-cortex-and-using-an-insula-searchlight), ‘sends out an alert’ to the CEN and Sensorimotor Networks as well as dampens the distracting activity of the DMN.
Professor: We’re out of time for this blog post, and the discussion of the next three stages (4 thru 6) will be in the next one:
- Stage 4 – “About 300–400 ms post-stimulus, neocortical regions, notably the premotor cortex and temporo-parietal areas, respond to the attentional shift with a signal that is indexed by the time-average P3b evoked potential.”
- Stage 5 – “The ACC also facilitates response selection and motor response via its links to the midcingulate cortex, supplementary motor cortex, and other motor areas “
- Stage 6 – Subjective report will include “Recognition and Discrimination,” plus “Scaling” shown in Davenport’s 2007 model, plus generation of a verbal report.
Take Home: This post is about the time course of symptom awareness: Within about .015 seconds, less than one breath, the Saliency Network (SN) of our brain can NOTICE something is strange. At the end of six stages of relatively fast cortical processing, for example, being aware of inspiratory load, we can in the last stage report being “short-of-breath”.
Next: Part 2 will include the last three processing stages also including discussion on generation of symptom report and common neural networks for DYSPNEA, PAIN, and STATIC HANDGRIP.
References
Cauda F, Torta DM-E, Sacco K, Geda E, D’Agata F, Costa T, Duca S, Geminiani G, Amanzio M. (2011) Functional connectivity of the insula in the resting brain. NeuroImage 55 (2011) 8–23
Davenport PW (2007) Chemical and mechanical loads: What have we learned? In: O’Donnell DE, Banzett RB, Carrieri-Kohlman V, Casaburi R, Davenport PW, Gandevia SC, Gelb AF, Mahler DA, Webb KA. (2007) Pathophysiology of Dyspnea in Chronic Obstructive Pulmonary Disease: A Roundtable. Proc Am Thorac Soc 4: 147–149 – See more at: http://www.endurance-education.com/endurance-education-com/the-lung-airways-are-connected-to-the-stretch-receptors-and-the-etc/#sthash.f7XNPBj7.dpuf
Menon, V. (2010) Large-scale brain networks in cognition: Emerging principles. In: Analysis and Function of Large-Scale Brain Networks. (Sporns O, ed) pp. 43-53. Washington, DC: Society for Neuroscience.
Raux, M., et al., (2013) Functional magnetic resonance imaging suggests automatization of the cortical response to inspiratory threshold loading in humans. Respir Physiol Neurobiol, http://dx.doi.org/10.1016/j.resp.2013.08.005
Sadaghiani S, Scheeringa R, Lehongre K, Morillon B, Giraud A-E, Kleinschmidt A. (2010) Intrinsic Connectivity Networks, Alpha Oscillations, and Tonic Alertness: A Simultaneous Electroencephalography / Functional Magnetic Resonance Imaging Study. DOI:10.1523/JNEUROSCI.1004-10.2010.
Sridharan D, Daniel J. Levitin DJ, Menon V. (2007) A critical role for the right fronto-insular cortex in switching between central-executive and default-mode networks. www.pnas.org/cgr/doi/10.1073/pnas.0800005105.
von Leopoldt A, Chan P-Y S, Esser RW, Davenport PW. (2013) Emotions and Neural Processing of Respiratory Sensations Investigated With Respiratory-Related Evoked Potentials. Psychosomatic Medicine 75: 244-252.
Weiser PC, Mahler DA, Ryan KP, Hill KL, Greenspon LW. (1993) Clinical assessment and management of dyspnea. In: Pulmonary rehabilitation: Guidelines to success. 2nd edition. Hodgkin, J, Bell CW, eds. Philadelphia: Lippincott, 478-511. – See more at: http://www.endurance-education.com/endurance-education-com/panting-starved-for-air-and-wheezing/#sthash.kzqTlbjq.dpuf