Eh! What? Motor programs operated by the brainstem?
Okay, my brainstem is the extension of my spinal cord. And I’m guessing this means just my brainstem selects each motor program I use?
Well, the short answer is Yes and No. Let’s allow the experts to explain!
First, in their paper entitled, ” Is there a brainstem substrate for action selection?”, Humphries et al. (2007) observe,
” Decerebrate animals and altricial (helpless at birth) neonates do not have fully intact basal ganglia but are capable of expressing spontaneous behaviours and coordinated and appropriate responses to stimuli. … Yet, the chronic decerebrate rat can, for example, spontaneously locomote, orient correctly to sounds, groom, perform coordinated feeding actions and discriminate food types …. Such animals clearly have some form of intact system for simple action selection that enables them to both respond to stimuli with appropriate actions (more complex than simple spinal-level reflexes), and sequence behaviours—as demonstrated by the holding, gnawing and chewing required for eating solid food.”
Grillner et al. (2013) have illustrated the vertebrate “motor infrastructure” in following figure:
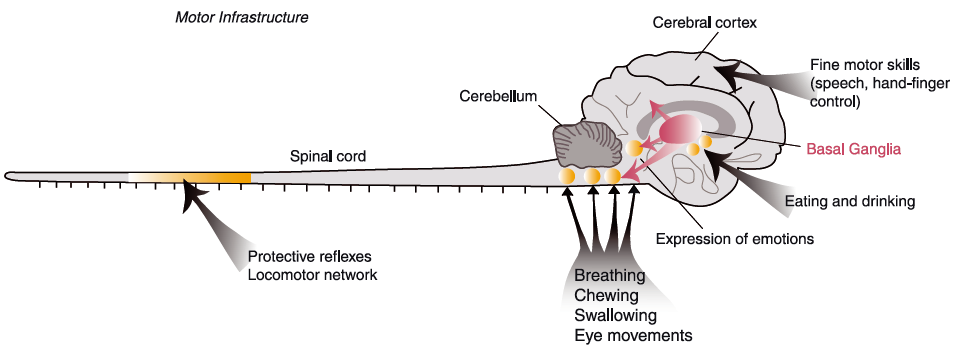
Figure 1. Common motor infrastructure from lamprey to man
Basic patterns of motor behaviour are controlled by neuronal networks (CPGs) located in the brainstem (e.g. swallowing, breathing) or the spinal cord (e.g. locomotion) (indicated as yellow circles), and the organisation is very similar throughout vertebrate phylogeny. The organisation of the basal ganglia is conserved from lamprey to primates. The basal ganglia control the activity in different brainstem motor centres and play a crucial role in the selection of motor behaviours. In primates and man a well-developed cerebral cortex provides a locus for networks controlling fine motor skills, and it also receives input from the basal ganglia via the thalamus.
In their very recent review, Grillner and coworkers (2013) also wrote that, “it may be important to note that mammals (e.g. cat) devoid of neocortex, but with all other parts of the forebrain intact, display basic goal-directed movements like searching for food and eating …, which is most likely dependent on the basal ganglia control of brainstem circuits.”
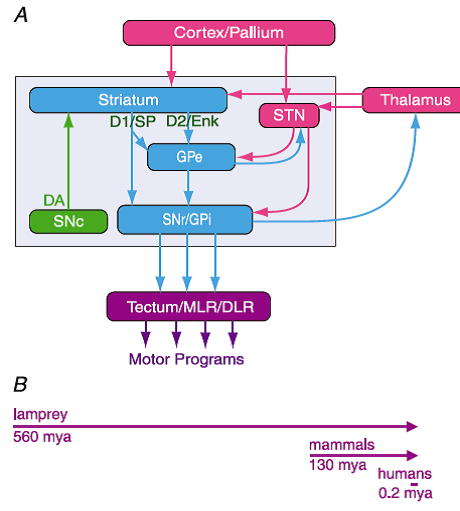
Figure 2. The organisation of the basal ganglia is almost identical throughout vertebrate phylogeny – from lamprey to primates
A, the striatum consists of GABAergic neurones (blue), as also globus pallidus externa (GPe), globus pallidus interna (GPi) and substantia nigra pars reticulata (SNr). SNr and GPi represent the output level of the basal ganglia, and it projects via different subpopulations of neurones to tectum (superior colliculus), the mesencephalic (MLR) and diencephalic (DLR) locomotor command regions and other brainstem motor centres, and also back to thalamus and cortex (pallium in lower vertebrates). The indirect loop is represented by the GPe, the subthalamic nucleus (STN) and the output level (SNr/GPi). The striatal neurones of the direct pathway to SNr/GPi express D1R and substance P (D1/SP), while the indirect pathway neurones in striatum express D2R and enkephalin (D2/Enk). Excitatory glutamatergic neurones are in pink. Also indicated is the dopamine supply from the substantia nigra pars compacta (SNc; green). B, the vertebrate lineage is represented. The lamprey had already diverged from the main vertebrate line 560 million years ago (mya); mammals emerged only some 130 mya and humans some 0.2 mya. Yet the design of the basal ganglia is conserved from lamprey to primates. In mammals there is a well-described pallido-thalamo-cortico projection that has not yet been investigated in the lamprey.
Please notice that the organization of the vertebrate basal ganglia consists of three parts.
- Motor output to the brainstem motor command areas (Tectum/MLR/DLR) come from only the substantia nigra pars reticulata (SNr) and globus pallidus interna (GPi).
NOTE: at rest, the neurons in these areas are under continuous strong inibition - Intrinsic direct and indirect pathways of the basal ganglia ultimately control the outputs of motor program selection. The activation of selection occur in the striatum, i.e., the caudate and putamen:
“The projection neurones in the striatum are of two types (see Fig. 2A), one of which projects directly to subpopulations of neurones at the output level (GPi, SNr) and express dopamine receptors of the D1 type (D1R), which mediate additional excitation. The other type of projection neurones express dopamine D2 receptors (D2R), which instead lower the excitability of this striatal neurone subtype that projects to globus pallidus externa (GPe), and this in turn interacts with the subthalamic nucleus (STN), which projects to the output level. The D1R projection neurones are referred as a direct pathway (to GPi and SNr) and are involved in the initiation of different motor programmes, while the D2R projection neurones are part of the indirect pathway, which inhibit movements and serve as a NO-GOpathway (deLong, 1990; Kravitz et al. 2010). There is also a ‘hyperdirect’ NO-GO pathway from cortex to the subthalamic nucleus that will enhance the inhibitory output.”
- Excitatory input from thalamus and cortex/pallium, first of all, acts on subpopulations of D1R projection neurons and thereby controls the basic aspects of motor selection. An example of input are prefrontal basal ganglia circuits. Another input pathway comes from the thalamus. The strength of these inputs determine whether or not the motor programs become activated or not. That is, these inputs have to dis-inhibit the tonic inhibition of the output neurons. Very interesting!!!!
For the purist scientist out there, there is a critical chunk of subcortical architecture that is missing: the pedunculopontine nucleus.
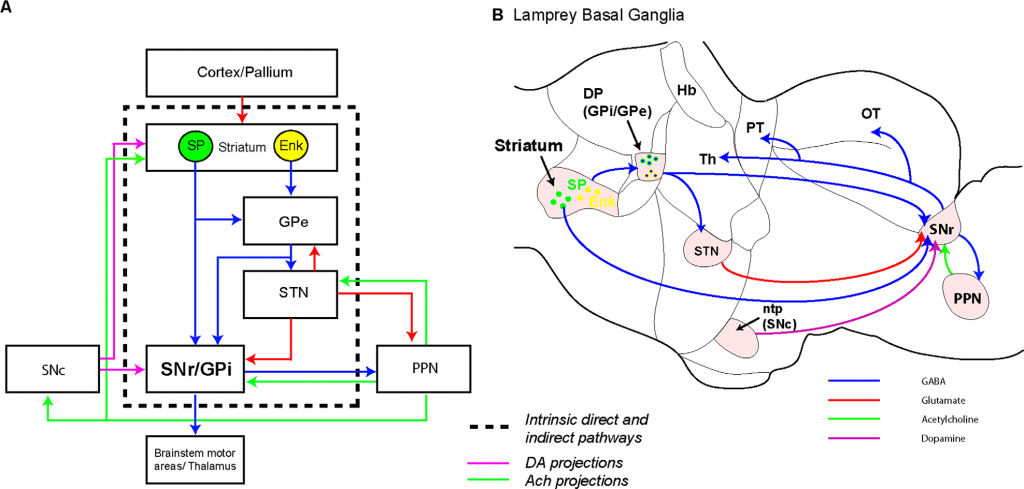
Figure 9. Summary of the organization of the substantia nigra pars reticulata and pedunculopontine nucleus in lamprey. A: Schematic showing the evolutionarily conserved architecture of the basal ganglia. Blue, red, green, and pink arrows indicate GABAergic, glutamatergic, cholinergic, and dopaminergic projections, respectively. B: Schematic sagittal section through the lamprey brain showing the location of the known basal ganglia nuclei and the connectivity of the SNr.
Figure 9 above found it in the Lamprey and completed the conserved basal ganglia-brainstem “core structure” according to Stephanson-Jones, Ericsson, Roberson, and Grillner (2012).
But what about the selection of specific motor programs?
Recent research findings actually suggest an anatomical basis for specificity. Check out the figure balow:
Figure 2. Anatomical organization of the vertebrate mRF. Directional arrows apply to both panels. (a) Sagittal section of the brainstem. The dendritic trees (thick grey lines) of the projection neurons (one neuron body shown, open circle) extend throughout the mRF along the dorsoventral axis but extend little along the rostrocaudal axis. These dendritic trees contact axon collaterals of both passing fibre systems (black dashed line) and far-reaching axons of the projection neurons (the axon of the depicted neuron body is shown by the black solid line); the example fibre system is the spinothalamic tract (ST). (b) The proposed mRF organization: it comprises stacked clusters (three of them are shown) containing medium-to-large projection neurons (open circles) and small-to-medium interneurons (filled circles); cluster limits (grey ovals) are defined by the initial collaterals from the projection neuron axons. The projection neurons’ radial dendritic fields allow sampling of ascending and descending inputs both from other clusters (solid black lines) and from passing fibre systems (dashed black line). The interneurons project within their parent cluster. Reproduced from Humphries et al. (2006).
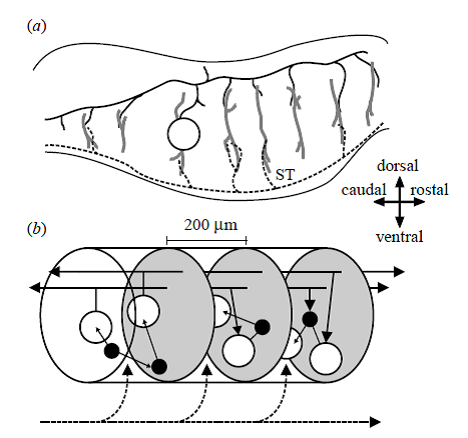
Please note the mRF [medial Reticular Formation] “Stacked Clusters” shown on Fig. 2 of the modeling paper by Humphries et al. (2007). Clusters are the “brainstem substrates” proposed by the investigators as the basis of the limited action selection in the limited functioning animals without basal ganglia and higher cortical centers.
Now look at the figure below by Grillner et al. (2013) for their proposed modular organization of the basal ganglia:
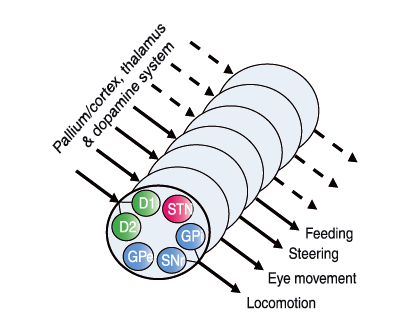
Figure 3. Conceptual scheme of a modular organisation of the basal ganglia, with one module for each type of motor programme
Each module would contain the D1R and D2R projection neurones and the components of the direct and indirect pathway GPi, SNr, GPe and STN (see Fig. 2A). Each module would be activated, if sufficient drive occurs from neurones in pallium/cortex and thalamus. The responsiveness of the modules would be determined by the tonic dopaminergic drive. Whereas the lamprey would have a limited behavioural repertoire and few modules, mammals and particularly primates have a very varied and versatile motor repertoire and presumably a greater number of modules.
Pleas note Grillner et al. (2013) comment on greater number of locomotor modules in the basal ganglia that could match up and include the stack clusters in Humphries and coworkers’ (2007) mLR stacked clusters.
In fact, Humphries et al. (2007) combined the selection schemes that feature the pedunculopontine nucleus as the critical structure allowing the feedforward-feedback action selection system to function properly:
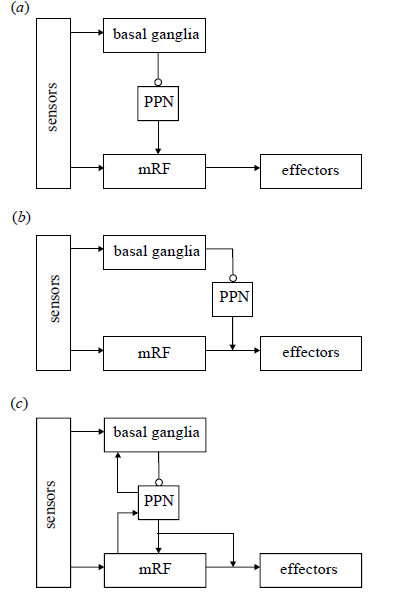
Figure 5. Alternative schemes for integrating the action selection substrates. (a) A hierarchical architecture: lower levels represent increasingly simple actions, selected by the higher layers. This is consistent with the output of the basal ganglia reaching the mRF via the PPN, and with the results of our modelling work. (b) A layered architecture: the mRF and basal ganglia form separate layers in a control system dealing with increasingly complex stimuli, the higher layers being able to veto the output of the lower layers. This design is consistent with the separate sensory input to the basal ganglia and mRF, and with the basal ganglia’s access to the spinal cord via the PPN. (c) A combined architecture: the competences of each layer contribute to the whole system. This is consistent with the evidence for feedback pathways within the neural systems, particularly between the PPN and the basal ganglia. Arrows, excitatory pathways; open circles, inhibitory pathways
Take home: Action selection involves a provocation to jolt the input striatal neurons that then activate basal ganglia neuron circuits to stimulate the specific module(s) for a specific motor program. The programs are as simple as eye movement or as complex as clearing the last steeplechase hurdle and dashing to the finish line.
Next: Is the basal ganglia involved in setting force produced by motor units and patterns? How do we prepare for and initiate locomotor tasks? And what exactly is the role of primary cortical motor area (M!)?
References
Grillner S, Robertson B, Stephenson-Jones M. (2013) The evolutionary origin of the vertebrate basal ganglia and its role in action selection. J Physiol doi: 10.1113/jphysiol.2012.246660.
Humphries MD, Gurney K, Prescott TJ. (2006) The brainstem reticular formation is a small-world, not scale-free, network. Phil Trans R Soc B 273: 503–511.
Humphries MD, Gurney K, Prescott TJ. (2007) Is there a brainstem substrate for action selection? Phil Trans R Soc B 362: 1627–1639
Stephenson-Jones M, Ericsson j, Robertson B, Grillner S. (2012) Evolution of the Basal Ganglia: Dual-Output Pathways Conserved Throughout Vertebrate Phylogeny. J Com. Neurol 520:2957–2973.